



スフィンゴ脂質が「曲がる」と細胞はどうなる?



~出芽酵母のスフィンゴ脂質を「シス二重結合含有型」へほぼ完全置換~
2025年8月8日
岐阜大学
スフィンゴ脂質が「曲がる」と細胞はどうなる? ~出芽酵母のスフィンゴ脂質を「シス二重結合含有型」へほぼ完全置換~
本研究のポイント
・出芽酵母のスフィンゴ脂質の基本骨格「スフィンゴイド塩基」を、植物型の構造へとほぼ完全に置き換えることに世界で初めて成功しました。
・スフィンゴ脂質の骨格構造の大部分にシス型二重結合が導入され、分子内に屈曲が生じても、出芽酵母は生育可能であることを示しました。
・スフィンゴ脂質がほぼ全てシス二重結合含有型になると、細胞膜全体の物性や脂質マイクロドメインの形成に異常が生じることを細胞レベルで示し、スフィンゴ脂質の「直鎖性」が細胞膜機能の維持に重要であることを実験的に示しました。
研究概要
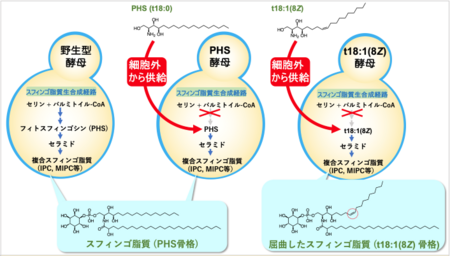
岐阜大学応用生物科学部応用生命化学科の谷 元洋教授らの研究グループは、分子遺伝学的手法を用いて、出芽酵母の生体膜を構成するスフィンゴ脂質の基本骨格構造 (スフィンゴイド塩基)を、植物に存在するシス二重結合含有型の(8Z)-4-ヒドロキシ-8-スフィンゲニン (t18:1(8Z))にほぼ完全に置き換えることに世界で初めて成功しました。
通常、細胞膜を構成するスフィンゴ脂質の疎水性部分の大部分は、直鎖状の炭化水素から成り、このような構造的特徴は細胞膜の秩序形成、機能維持に重要であると考えられてきました。本研究では、ほぼすべてのスフィンゴ脂質分子に屈曲構造(シス二重結合)を導入した出芽酵母を構築し、細胞膜構造や脂質マイクロドメイン形成に生じる変化を詳細に解析しました。その結果、スフィンゴ脂質の「直鎖性」が細胞膜の正常な機能維持に不可欠であることを、細胞レベルで初めて実験的に示しました。今後、このような出芽酵母を膜脂質機能解明のためのモデルプラットフォームとしてさらに活用することで、「生体膜脂質がなぜ多様な構造を持つのか?」という生命科学の根本的問いに迫れることが期待されます。
本研究成果は、現地時間2025年8月5日に欧州生化学連合の国際誌FEBS Open Bioのオンライン版に掲載されました。
研究背景
スフィンゴ脂質注1)は、真核細胞の膜構成成分として不可欠な脂質であり、細胞内外の情報伝達、物質輸送、ストレス応答などに関与しています。スフィンゴ脂質は、①スフィンゴイド塩基、②脂肪酸、③親水性頭部の3つの要素で構成されており、これらの構造ユニットの多様性により、スフィンゴ脂質は複雑な構造を持っています (図1)。
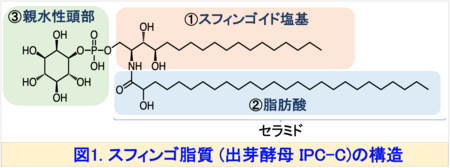
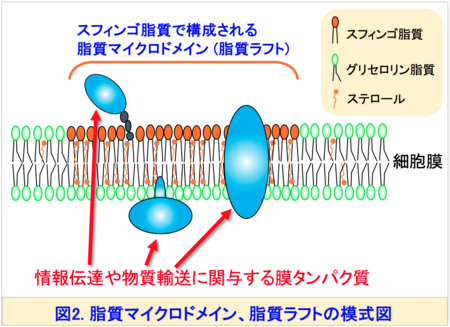
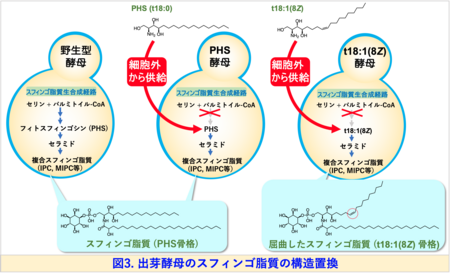
この多様性はスフィンゴ脂質が多彩な生理機能を果たすための分子基盤と考えられています。また、スフィンゴ脂質の構造は生物種間で明確に異なることがあります。たとえば、真核生物の最も単純なモデル生物である出芽酵母注2)では、スフィンゴイド塩基のほとんどが二重結合を持たないフィトスフィンゴシン (PHS)ですが、植物のスフィンゴイド塩基の一部には、シス二重結合を持つ構造 ((8Z)-4-ヒドロキシ-8-スフィンゲニン (t18:1(8Z)) (図3右上)が存在します。一般的にスフィンゴ脂質は、脂質ラフトあるいは脂質マイクロドメイン注3)と呼ばれる秩序性の高いドメイン構造を細胞膜中で形成します。これらの形成には、疎水性部分 (セラミド) (図1)が膜中で直鎖状に密集して存在し、秩序立った構造をつくることが鍵となると考えられています (図2)。したがって、シス二重結合によって屈曲したスフィンゴ脂質の存在意義は、これまで十分に解明されていませんでした。
研究成果
本研究では、出芽酵母のスフィンゴ脂質の生合成の最初の代謝ステップに関与する酵素遺伝子を破壊し、細胞外から任意のスフィンゴイド塩基を供給することで、スフィンゴ脂質の骨格構造のほぼすべてをt18:1(8Z)に置き換えた変異酵母 (t18:1(8Z)酵母)を樹立することに成功しました (図3)。
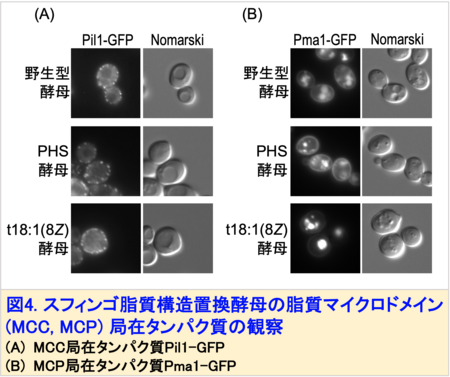
この変異酵母は、野生型酵母もしくはPHSを供給した変異酵母(PHS酵母)に比べて生育速度がやや低下するものの、外見上の大きな変化は観察されませんでした。しかしながら、t18:1(8Z)酵母は細胞膜の透過性上昇や秩序性低下が観察され、細胞膜が脆弱化していることが示唆されました。出芽酵母には、脂質マイクロドメインとして、MCC (Membrane Compartment of Can1)、MCP (Membrane Compartment of Pma1)と呼ばれる構造があります。そこで、MCCに局在するタンパク質Pil1, MCPに局在するタンパク質Pma1にGFP注4)を付加した融合タンパク質を発現させ、細胞内での局在を観察しました。その結果、Pil1-GFPは細胞膜上に点状に分布しますが、t18:1(8Z)酵母ではその点の数が増加していました (図4A)。さらにPma1-GFPは、細胞膜に留まることができず、そのほとんどが液胞に蓄積することがわかりました (図4B)。
このようにスフィンゴ脂質が屈曲することで、細胞膜全体の物性や脂質マイクロドメインの形成に異常が生じることが明らかになりました。興味深いことに、t18:1(8Z)酵母では、細胞壁注5)の強度が上昇していることも確認されました。これは、細胞膜の脆弱化に対して細胞壁を強化するという“補償的な適応反応”が誘導されることを示唆しています。
以上の結果より、スフィンゴ脂質の骨格が屈曲構造を持つことにより、細胞膜の構造的完全性(インテグリティ)や、脂質マイクロドメインの形成に重大な影響が及ぶことが明らかとなりました。これらのことから、スフィンゴ脂質の「直鎖性」は、脂質マイクロドメインを含む細胞膜の秩序形成と機能維持にとって本質的な構造特性であるという従来の考えを、細胞レベルで実験的に示すことができました。
今後の展開
膜脂質は、すべての細胞において、外界と細胞内を区画化する膜構造の形成に不可欠な分子です。自然界には数万種類の膜脂質が存在すると言われています。仮に膜脂質が単に区画を形成する「壁」としての役割にとどまるのであれば、これほどまでの構造多様性は必要ないはずです。したがって、膜脂質構造多様性の生物学的意義の解明は生命科学における重要な課題といえます。本研究で行ったように、出芽酵母の膜脂質を異種生物の構造に完全に入れ替える「構造置換」手法は、この構造多様性の意義を明らかにするための有力なアプローチとなると考えます。今後は、スフィンゴ脂質に関して、スフィンゴイド塩基だけでなく、親水性頭部や脂肪酸部分(図1)についても構造置換を組み合わせることで、出芽酵母を膜脂質機能解明のためのモデルプラットフォームとしてさらに発展させていきたいと考えています。
用語解説
注1) スフィンゴ脂質
真核生物の細胞膜を構成する主要な脂質の一つで、細胞の構造保持や情報伝達に重要な役割を果たしている。
注2) 出芽酵母
パンや酒の発酵に使われる単細胞の真核生物で、細胞の基本的な仕組みを研究するモデル生物として広く利用されている。
注3) 脂質マイクロドメイン (脂質ラフト)
細胞膜内に存在する特定の脂質やタンパク質が集まってできる「島状」の構造で、シグナル伝達や膜タンパク質の機能に関与する。
注4) GFP
緑色蛍光タンパク質。特定のタンパク質と融合させることで、生きた細胞内でのその動きや局在を可視化できる。
注5) 細胞壁
酵母や植物の細胞を外側から包む丈夫な構造で、細胞の形を保ち、外部からの様々なストレスに耐える役割を担う。
論文情報
雑誌名:FEBS Open Bio
論文タイトル:Cis-unsaturated sphingolipids support growth of sphingoid base-deficient yeast but impair plasma membrane integrity
著者:Takashi Higuchi¶, Saki Sugihara¶, Yohei Ishibashi, Kono Yushi, Hazuki Yamauchi, and Motohiro Tani* ¶: equal contribution *: corresponding author
DOI: 10.1002/2211-5463.70100
研究支援
本研究は科学研究費助成事業(科研費)の挑戦的研究(萌芽) 23K18009 及び基盤研究(B) 24K01682 の支援を受けました。
-
出芽酵母のスフィンゴ脂質の構造置換 -
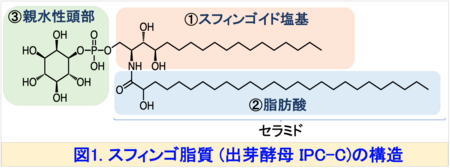
図1 -
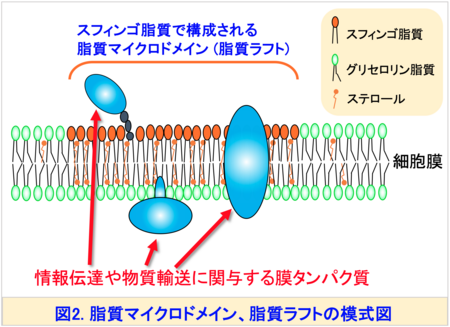
図2 -
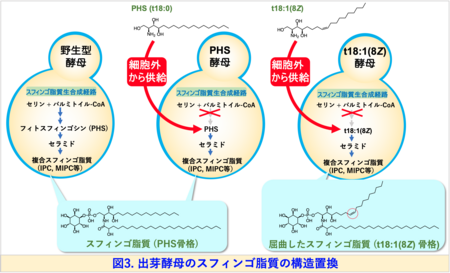
図3 -
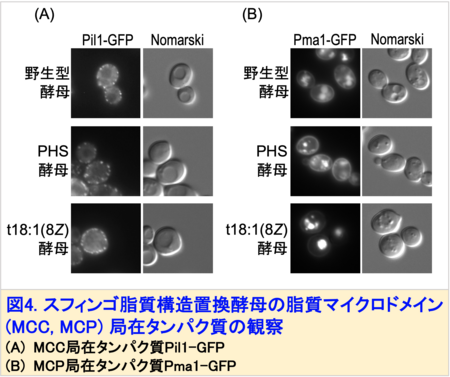
図4















